
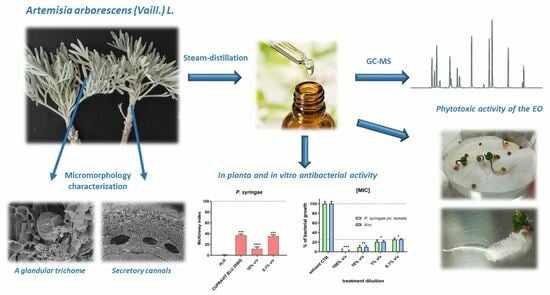
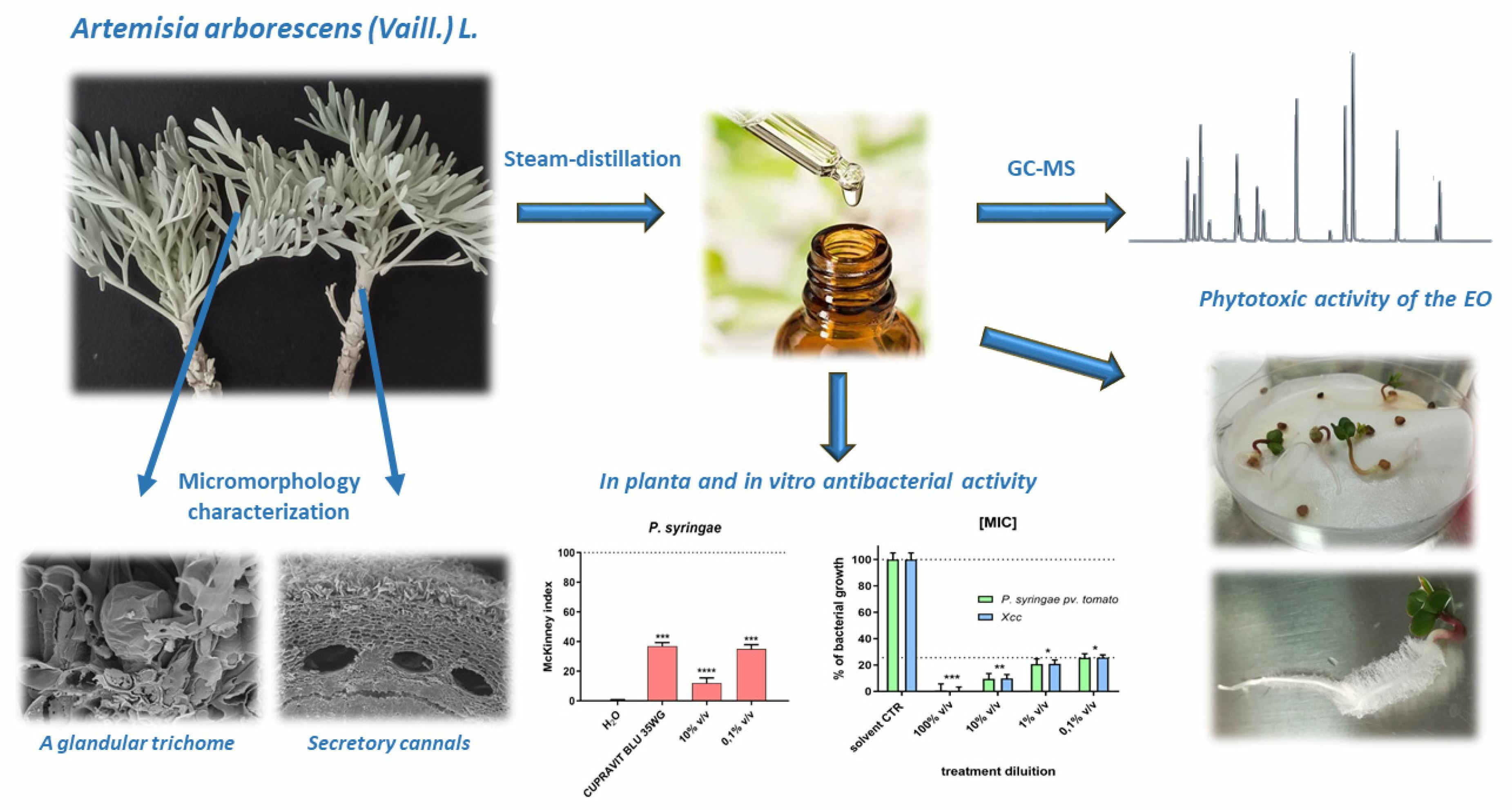
Artemisia arborescens (Vaill.) L.: Micromorphology, Essential Oil Composition, and Its Potential as an Alternative Biocontrol Product
 , , , and
, , , and
Abstract
:
1. Introduction
2. Results
2.1. Anatomical and Micromorphological Analyses
2.1.1. Leaves
2.1.2. Petiole and Young Stem
2.2. Chemical Composition of EO
2.3. Phytotoxic Activity
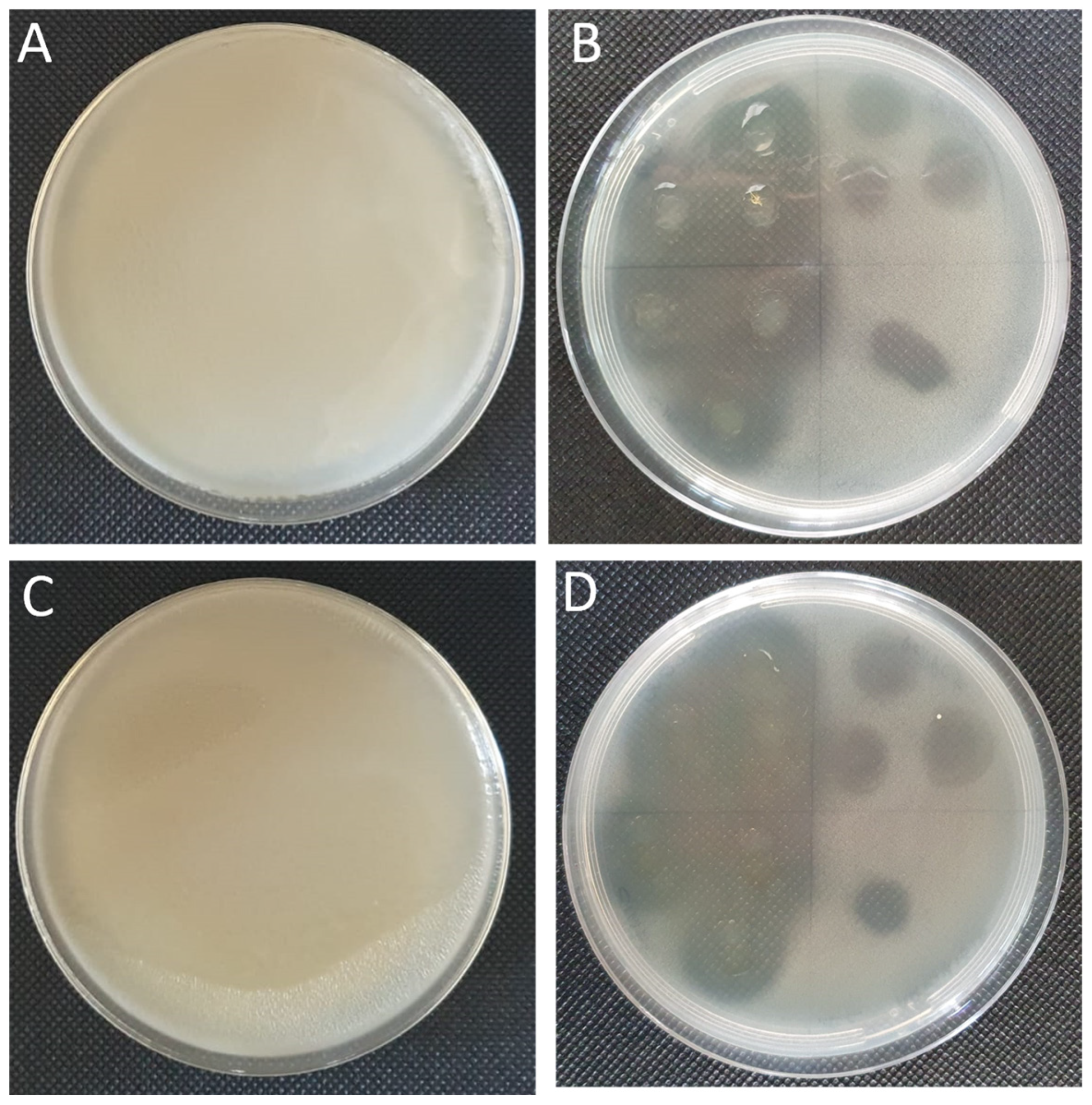
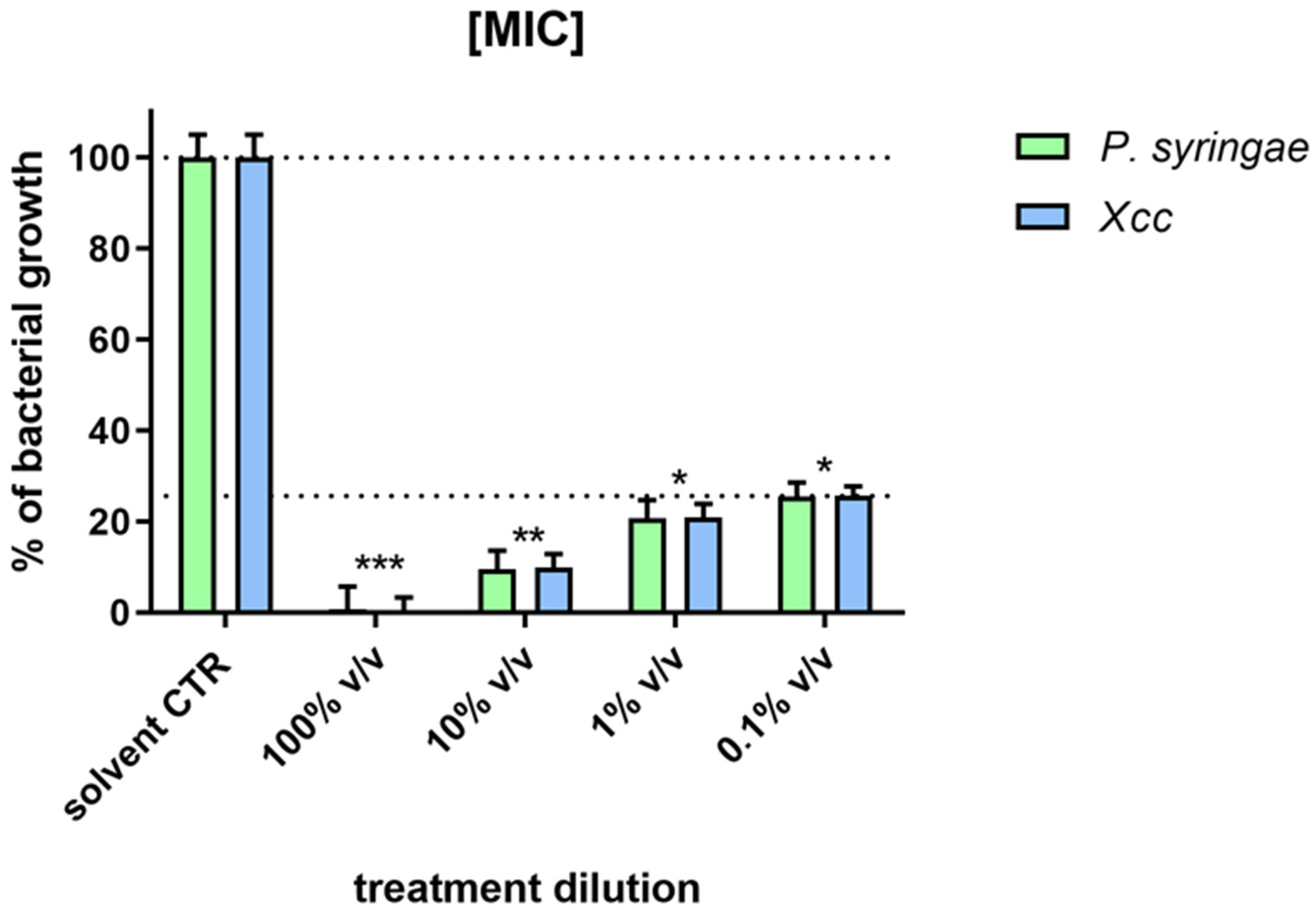
2.4. Antimicrobial and Antibiofilm Activity of the Essential Oil
2.5. Activity of EO in Planta
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Light and Scanning Electron Microscopy
4.3. Extraction of Essential Oils
4.4. Analysis of Essential Oils
4.5. Phytotoxic Activity
4.6. Plant Pathogens
4.7. Antimicrobial Activity
4.8. Antibiofilm Activity
4.9. Bioassay on Tomato Leaves Infected with Pseudomonas syringae pv. tomato
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bora, K.S.; Sharma, A. The genus Artemisia: A comprehensive review. Pharm. Biol. 2011, 49, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Nigam, M.; Atanassova, M.; Mishra, A.P.; Pezzani, R.; Devkota, H.P.; Plygun, S.; Salehi, B.; Setzer, W.N.; Sharifi-Rad, J. Bioactive Compounds and Health Benefits of Artemisia Species. Nat. Prod. Commun. 2019, 14, 7. [Google Scholar] [CrossRef]
- Lai, F.; Loy, G.; Manconi, M.; Manca, M.L.; Fadda, A.M. Artemisia arborescens L. essential oil loaded beads: Preparation and characterization. AAPS PharmSciTech 2007, 8, E126–E132. [Google Scholar] [CrossRef] [PubMed]
- Atzei, A.D. Le Piante Nella Tradizione Popolare della Sardegna; Delfino Editore: Sassari, Italy, 2003. [Google Scholar]
- Guarrera, P.M. Usi e Tradizioni della Flora Italiana; Aracne Editrice: Roma, Italy, 2006. [Google Scholar]
- Michelakis, E.C.; Evergetis, E.; Koulocheri, S.D.; Haroutounian, S.A. Exploitation of Artemisia arborescens as a renewable source of chamazulene: Seasonal variation and distillation conditions. Nat. Prod. Commun. 2016, 11, 1513–1516. [Google Scholar] [CrossRef]
- Abu Zarga, M.; Qauasmeh, R.; Sabri, S.; Munsoor, M.; Abdalla, S. Chemical constituents of Artemisia arborescens and the effect of the aqueous extract on rat isolated smooth muscle. Planta Med. 1995, 61, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Creed, C.; Mollhagen, A.; Mollhagen, N.; Pszczolkowski, M.A. Artemisia arborescens “Powis Castle” extracts and α-thujone prevent fruit infestation by codling moth neonates. Pharm. Biol. 2015, 53, 1458–1464. [Google Scholar] [CrossRef] [PubMed]
- Militello, M.; Settanni, L.; Aleo, A.; Mammina, C.; Moschetti, G.; Giammanco, G.M.; Blázquez, M.D.; Carrulba, A. Chemical composition and antibacterial potential of Artemisia arborescens L. essential oil. Curr. Microbiol. 2011, 62, 1274–1281. [Google Scholar] [CrossRef]
- Saddi, M.; Sanna, A.; Bottiglia, F.; Chisu, L.; Casu, L.; Bonsignore, L.; De Laguna, A. Anti herpesvirus activity of Artemisia arborescens essential oil and inhibition of lateral diffusion in Vero cells. Ann. Clin. Microbiol. Antimicrob. 2007, 6, 10–16. [Google Scholar] [CrossRef]
- Lydon, J.; Teasdale, J.R.; Chen, P.K. Allelopathic Activity of Annual Wormwood (Artemisia annua) and the Role of Artemisinin. Weed Sci. 1997, 45, 807–811. [Google Scholar] [CrossRef]
- Li, J.; Chen, L.; Chen, Q.; Miao, Y.; Peng, Z.; Huang, B.; Guo, L.; Liu, D.; Du, H. Allelopathic effect of Artemisia argyi on the germination and growth of various weeds. Sci. Rep. 2021, 11, 4304. [Google Scholar] [CrossRef]
- Li, J.; Zhao, T.; Chen, L.; Hong, C.; Luo, D.; Chen, C.; Miao, Y.; Liu, D. Artemisia argyi allelopathy: A generalist compromises hormone balance, element absorption, and photosynthesis of receptor plants. BMC Plant Biol. 2022, 22, 368. [Google Scholar] [CrossRef]
- Araniti, F.; Lupini, A.; Sorgonà, A.; Conforti, F.; Marrelli, M.; Statti, G.A.; Menichini, F.; Abenavoli, M.R. Allelopathic potential of Artemisia arborescens: Isolation, identification and quantification of phytotoxic compounds through fractionation-guided bioassays. Nat. Prod. Res. 2013, 27, 880–887. [Google Scholar] [CrossRef]
- Labruzzo, A.; Cantrell, C.L.; Carrubba, A.; Ali, A.; Wedge, D.E.; Duke, S.O. Phytotoxic Lignans from Artemisia arborescens. Nat. Prod. Commun. 2018, 13, 237–240. [Google Scholar] [CrossRef]
- Ilehi, O.; Chograni, H.; Rejeb, F.B.; Riahi, L. Selectivity of the bioherbicidal potential of Artemisia arborescens L. among model and agronomic crops and implications for pre- and post-emergence applications. Bot. Sci. 2023, 101, 1128–1144. [Google Scholar] [CrossRef]
- Araniti, F.; Gullì, T.; Marrelli, M.; Statti, G.; Gelsomino, A.; Abenavoli, M.R. Artemisia arborescens L. leaf litter: Phytotoxic activity and phytochemical characterization. Acta Physiol. Plant. 2016, 38, 128. [Google Scholar] [CrossRef]
- Dudai, A.N.D.; Poljakoff-Maybe, A.; Mayer, A.M.; Putievsky, E.; Lerner, H.R. Essential oils as allelochemicals and their potential use as bioherbicides. J. Chem. Ecol. 1999, 25, 1079–1089. [Google Scholar] [CrossRef]
- Hazell, P.; Wood, S. Drivers of change in global agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2007, 363, 495–515. [Google Scholar] [CrossRef] [PubMed]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed]
- Rauscher, C.I.E.D.E.M.; Rauscher, M. International Trade, FactorMovements, and the Environment; Clarendon Press: Oxford, UK, 1997; ISBN 9780198290506. [Google Scholar]
- Pilet-Nayel, M.L.; Moury, B.; Caffier, V.; Montarry, J.; Kerlan, M.C.; Fournet, S.; Durel, C.E.; Delourme, R. Quantitative resistance to plant pathogens in pyramiding strategies for durable crop protection. Front. Plant Sci. 2017, 8, 1838. [Google Scholar] [CrossRef]
- Usta, C. Microorganisms in Biological Pest Control—A Review (Bacterial Toxin Application and Effect of Environmental Factors). In Current Progress in Biological Research; InTech Open: London, UK, 2013; Volume 32, pp. 137–144. [Google Scholar]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Macfadyen, S.; Hardie, D.C.; Fagan, L.; Stefanova, K.; Perry, K.D.; DeGraaf, H.E.; Holloway, J.; Spafford, H.; Umina, P.A. Reducing Insecticide Use in Broad-Acre Grains Production: An Australian Study. PLoS ONE 2014, 9, e89119. [Google Scholar] [CrossRef] [PubMed]
- Tworkoski, T. Herbicide effects of essential oils. Weed Sci. 2002, 50, 425–431. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends. Plant. Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.R.; Santomauro, F.; Sacco, C.; Bergonzi, M.C.; Donato, R. Essential Oil of Artemisia annua L.: An Extraordinary Component with Numerous Antimicrobial Properties. Evid.-Based Complement. Altern. Med. 2014, 2014, 159819. [Google Scholar] [CrossRef]
- Bartoli, A.; Galati, B.G.; Tortosa, R.D. Anatomical studies of the secretory structures: Glandular trichomes and ducts, in Grindelia pulchella Dunal (Astereae, Asteraceae). Flora 2011, 206, 1063–1068. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure–function coordination. J. For. Res. 2020, 31, 1–12. [Google Scholar] [CrossRef]
- Harbone, J.B. Introduction to Ecological Biochemistry; Academic Press: London, UK, 1993. [Google Scholar]
- Dussourd, D.E.; Denno, R.F. Deactivation of plant defense: Correspondence between insect behavior and secretory canal architecture. Ecology 1991, 72, 1383–1396. [Google Scholar] [CrossRef]
- Fahn, A. Functions and location of secretory tissues in plants and their possible evolutionary trends. Isr. J. Plant Sci. 2002, 50, 59–64. [Google Scholar] [CrossRef]
- Ascensão, L.; Pais, M.S.S. Glandular Trichomes of Artemisia campestris (ssp. maritima): Ontogeny and Histochemistry of the Secretory Product. Bot. Gaz. 1987, 148, 221–227. [Google Scholar] [CrossRef]
- Ascensão, L.; Pais, M.S. Ultrastructure and histochemistry of secretory ducts in Artemisia campestris ssp. maritima (Compositae). Nord. J. Bot. 1988, 8, 283–292. [Google Scholar] [CrossRef]
- Pagni, A.M.; Orlando, R.; Masini, A.; Ciccarelli, D. Secretory structures of Santolina ligustica Arrigoni (Asteraceae), an Italian endemic species. Isr. J. Plant. Sci. 2003, 51, 185–192. [Google Scholar] [CrossRef]
- Andreucci, A.C.; Ciccarelli, D.; Desideri, I.; Pagni, A.M. Glandular hairs and secretory ducts of Matricaria chamomilla (Asteraceae): Morphology and histochemistry. Ann. Bot. Fenn. 2008, 45, 11–18. [Google Scholar] [CrossRef]
- Lusa, M.G.; Da Costa, F.B.; Appezzato-da-Glória, B. Histolocalization of chemotaxonomic markers in Brazilian Vernonieae (Asteraceae). Bot. J. Linn. Soc. 2016, 182, 581–593. [Google Scholar] [CrossRef]
- Kelsey, R.G.; Shafizadeh, F. Glandular trichomes and sesquiterpene lactones of Artemisia nova (Asteraceae). Biochem. Syst. Ecol. 1980, 8, 371–377. [Google Scholar] [CrossRef]
- Cappelletti, E.M.; Caniato, R.; Appendino, G. Localization of the cytotoxic hydroperoxyeudesmanolides in Artemisia umbelliformis. Biochem. Syst. Ecol. 1986, 14, 183–190. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Mendes, A.R.; Caeiro, M.F.; Figueiredo, A.C.; Ascensão, L. New Reports on the Portuguese Endemic Species, Santolina impressa: Secretory Structures, Essential Oil Composition and Antiviral Activity. Plants 2023, 12, 2391. [Google Scholar] [CrossRef]
- Duke, S.O.; Paul, R.N. Development and Fine Structure of the Glandular Trichomes of Artemisia annua L. Int. J. Plant Sci. 1993, 154, 107–118. [Google Scholar] [CrossRef]
- Janaćković, P.; Gavrilović, M.; Rančić, D.; Dajić-Stevanović, A.A.; Marin, P. Comparative anatomical investigation of five Artemisia L. (Anthemideae, Asteraceae) species in view of taxonomy. Braz. J. Bot. 2019, 42, 135–147. [Google Scholar] [CrossRef]
- Ornano, L.; Venditti, A.; Ballero, M.; Sanna, C.; Quassinti, L.; Bramucci, M.; Lupidi, G.; Pope, F.; Vittori, S.; Maggi, F.; et al. Chemopreventive and antioxidant activity of the chamazulene-rich essential oil obtained from Artemisia arborescens L. growing on the Isle of La Maddalena, Sardinia, Italy. Chem. Biodivers. 2013, 10, 1464–1474. [Google Scholar] [CrossRef]
- Ghasemi, M.; Babaeian Jelodar, N.; Modarresi, M.; Bagheri, N.; Jamali, A. Increase of Chamazulene and α-Bisabolol contents of the essential oil of German Chamomile (Matricaria chamomilla L.) using salicylic acid treatments under normal and heat stress conditions. Foods 2016, 5, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Presti, M.L.; Crupi, M.L.; Zellner, B.D.A.; Dugo, G.; Mondello, L.; Dugo, P.; Ragusa, S. Characterization of Artemisia arborescens L. (Asteraceae) leaf-derived essential oil from Southern Italy. J. Essent. Oil Res. 2007, 19, 218–224. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis: A fundamental concept in biology. Microb. Cell. 2014, 1, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Naeem Mushtaq, M.; Yousa, M.; Sarwar, N. Allelopathic hormesis and potent allelochemicals from multipurpose tree Moringa oleifera leaf extract. Plant Biosyst. 2021, 155, 154–158. [Google Scholar] [CrossRef]
- Saad, Y.; Muller, D.; Lobstein, A. Major bioactivities and mechanism of action of essential oils and their components. Flavour Fragr. J. 2013, 28, 269–279. [Google Scholar] [CrossRef]
- Singh, N.B.; Devi, M.L.; Biona, T.; Sharma, N.; Das, S.; Chakravorty, J.; Mukherjee, P.K.; Rajashekar, Y. Phytochemical Composition and Antimicrobial Activity of Essential Oil from the Leaves of Artemisia vulgaris L. Molecules 2023, 28, 2279. [Google Scholar] [CrossRef] [PubMed]
- Adorjan, B.; Buchbauer, G. Biological properties of essential oils: An updated review. Flavour Fragr. J. 2010, 25, 407–426. [Google Scholar] [CrossRef]
- Das, S.; Vörös-Horváth, B.; Bencsik, T.; Micalizzi, G.; Mondello, L.; Horváth, G.; Kőszegi, T.; Széchenyi, A. Antimicrobial Activity of Different Artemisia Essential Oil Formulations. Molecules 2020, 25, 2390. [Google Scholar] [CrossRef]
- Kotan, R.; Kordali, S.; Cakir, A. Screening of antibacterial activities of twenty-one oxygenated monoterpenes. Z. Naturforsch. C. J. Biosci. 2007, 62, 507–513. [Google Scholar] [CrossRef]
- Noriega, P.; Ballesteros, J.; De la Cruz, A.; Veloz, T.X. Chemical Composition and Preliminary Antimicrobial Activity of the Hydroxylated Sesquiterpenes in the Essential Oil from Piper barbatum Kunth Leaves. Plants 2019, 9, 211. [Google Scholar] [CrossRef]
- Gómez-Sequeda, N.; Cáceres, M.; Stashenko, E.E.; Hidalgo, W.; Ortiz, C. Antimicrobial and Antibiofilm Activities of Essential Oils against Escherichia coli O157:H7 and Methicillin-Resistant Staphylococcus aureus (MRSA). Antibiotics 2020, 9, 730. [Google Scholar] [CrossRef]
- Jackson, B.P.; Snowdon, D.W. Atlas of Microscopy of Medicinal Plants, Culinary Herbs and Spices; Belhaven Press: London, UK, 1990. [Google Scholar]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with sudan red 7B or fluorol yellow 088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Chieco, C.; Rotondi, A.; Morrone, L.; Rapparini, F.; Baraldi, R. An ethanol-based fixation method for anatomical and micro-501 morphological characterization of leaves of various tree species. Biotech. Histochem. 2013, 88, 109–119. [Google Scholar] [CrossRef]
- O’Brien, T.P.; Feder, N.; Mccully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 9, 368–373. [Google Scholar] [CrossRef]
- O’Brien, T.P.; McCully, M.E. The Study of Plant Structure: Principles and Selected Methods; Termocarphy: Melbourne, Australia, 1981. [Google Scholar]
- Council of Europe. European Pharmacopoeia, 10th ed.; Council of Europe: Strasbourg, France, 2020. [Google Scholar]
- Jennings, W.; Shibamoto, T. Qualitative Analyisis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicone and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 1998. [Google Scholar]
- Goodner, K.I. Practical retention index models of OV-101, DB-1, DB-5, and DB-Wax for flavour and fragrance compounds. LWT-Food Sci. Technol. 2008, 41, 951–958. [Google Scholar] [CrossRef]
- McLafferty, F.W. The Wiley Registry of Mass Spectral Data, with Nist Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1998. [Google Scholar]
- Danna, C.; Cornara, L.; Smeriglio, A.; Trombetta, D.; Amato, G.; Aicardi, P.; De Martino, L.; De Feo, V.; Caputo, L. Eucalyptus gunnii and Eucalyptus pulverulenta ‘Baby Blue’ Essential Oils as Potential Natural Herbicides. Molecules 2021, 26, 6749. [Google Scholar] [CrossRef]
- Malaspina, P.; Papaianni, M.; Ranesi, M.; Polito, F.; Danna, C.; Aicardi, P.; Cornara, L.; Woo, S.L.; De Feo, V. Eucalyptus cinerea and E. nicholii by-Products as Source of Bioactive Compounds for Agricultural Applications. Plants 2022, 11, 2777. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed Germination and Dormancy. Plant. Cell. 1997, 9, 1055–1066. [Google Scholar] [CrossRef]
- Garbe, J.; Wesche, A.; Bunk, B.; Kazmierczak, M.; Selezska, K.; Rohde, C.; Jahn, D.; Schobert, M. Characterization of JG024, a Pseudomonas aeruginosa PB1-like broad host range phage under simulated infection conditions. BMC Microbiol. 2010, 10, 301. [Google Scholar] [CrossRef]
- Papaianni, M.; Ricciardelli, A.; Casillo, A.; Corsaro, M.M.; Borbone, F.; Della Ventura, B.; Velotta, R.; Fulgione, A.; Woo, S.L.; Tutino, M.L.; et al. The Union Is Strength: The Synergic Action of Long Fatty Acids and a Bacteriophage against Xanthomonas campestris Biofilm. Microorganisms 2021, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- McKinney, H.H. Influence of soil temperature and moisture on infection of wheat seedlings by Helmintosporium sativum. J. Agric. Res. 1925, 26, 195–218. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KI a | KI b | % | Identification c | |
|---|---|---|---|---|
| Tricyclene | 844 | 1047 | 0.3 | 1, 2 |
| α-Thujene | 850 | 1020 | 0.1 | 1, 2 |
| α-Pinene | 855 | 1036 | 2.3 | 1, 2, 3 |
| Camphene | 868 | 1075 | 6.0 | 1, 2, 3 |
| Butanoic acid, 2-methylpropyl ester | 870 | 1174 | 0.1 | 1, 2 |
| Sabinene | 891 | 1115 | 2.4 | 1, 2 |
| β-Pinene | 910 | 1120 | 0.8 | 1, 2, 3 |
| α-Phellandrene | 918 | 1177 | 1.8 | 1, 2, 3 |
| α-Terpinene | 929 | 1170 | 0.6 | 1, 2, 3 |
| Propanoic acid, 2-methyl-, 3-methylbutyl ester | 934 | 0.2 | 1, 2 | |
| p-Cymene | 938 | 1250 | 2.4 | 1, 2, 3 |
| Eucalyptol | 942 | 1210 | 2.2 | 1, 2, 3 |
| γ-Terpinene | 970 | 1221 | 1.0 | 1, 2, 3 |
| Terpinolene | 997 | 1267 | 0.2 | 1, 2, 3 |
| cis-Thujone | 1009 | 1430 | 1.2 | 1, 2 |
| trans-Thujone | 1020 | 1442 | 24.2 | 1, 2 |
| α-Campholenal | 1032 | 1485 | 0.2 | 1, 2 |
| allo-Ocimene | 1041 | 1388 | 0.1 | 1, 2 |
| Camphor | 1050 | 1491 | 18.9 | 1, 2, 3 |
| Pinocarvone | 1063 | 1586 | 0.1 | 1, 2, 3 |
| Borneol | 1067 | 1715 | 0.2 | 1, 2, 3 |
| Terpinen-4-ol | 1079 | 1590 | 0.7 | 1, 2, 3 |
| Isocitral | 1084 | 1690 | 0.1 | 1, 2 |
| α-Terpineol | 1093 | 1661 | 0.3 | 1, 2 |
| cis-Chrysantenyl acetate | 1098 | 0.2 | 1, 2 | |
| 4-Decen-1-ol | 1133 | 0.2 | 1, 2 | |
| Thymol | 1199 | 2172 | 0.6 | 1, 2, 3 |
| δ-Elemene | 1219 | 1479 | 0.1 | 1, 2 |
| α-Copaene | 1255 | 1477 | 0.4 | 1, 2 |
| Isobornyl propanoate | 1258 | 0.1 | 1, 2 | |
| β-Bourbonene | 1263 | 1498 | 0.3 | 1, 2 |
| β-Elemene | 1266 | 1579 | 0.1 | 1, 2 |
| β-Gurjunene | 1273 | 1615 | 1.2 | 1, 2 |
| Aromadendrene | 1298 | 1631 | 6.5 | 1, 2 |
| cis-Muurola-3,5-diene | 1299 | 0.7 | 1, 2 | |
| α-Humulene | 1322 | 1641 | 0.8 | 1, 2 |
| γ-Gurjunene | 1346 | 0.7 | 1, 2 | |
| Germacrene D | 1350 | 1712 | 1.5 | 1, 2 |
| cis-β-Guaiene | 1354 | 1651 | 0.2 | 1, 2 |
| trans-Muurola-4(14),5-diene | 1365 | 0.3 | 1, 2 | |
| γ-Amorphene | 1383 | 0.5 | 1, 2 | |
| δ-Amorphene | 1393 | 1751 | 0.5 | 1, 2 |
| α-Cadinene | 1400 | 1753 | 0.1 | 1, 2 |
| Caryophyllene oxide | 1443 | 2000 | 3.0 | 1, 2 |
| Aristolene epoxide | 1453 | 0.2 | 1, 2 | |
| Caryophylla-4(12),8(13)-dien-5α-ol | 1498 | 2324 | 0.8 | 1, 2 |
| α-Cadinol | 1507 | 2256 | 1.3 | 1, 2 |
| Guaia-3,10(14)-dien-11-ol | 1538 | 0.1 | 1, 2 | |
| cis-Z-α-Bisabolene epoxide | 1550 | 0.1 | 1, 2 | |
| Cedren-13-ol, 8- | 1837 | 2359 | 5.2 | 1, 2 |
| trans-Nuciferol | 1839 | 4.2 | 1, 2 | |
| Total | 96.3 | |||
| Monoterpene hydrocarbons | 18.0 | |||
| Oxygenated monoterpenes | 49.0 | |||
| Sesquiterpene hydrocarbons | 13.9 | |||
| Oxygenated sesquiterpenes | 14.9 | |||
| Others | 0.5 |
| Number of Germinated Seeds (A) and Percent Inhibition (B) | ||||||||
|---|---|---|---|---|---|---|---|---|
| µg/mL | L. multiflorum | S. arvensis | R. sativus | C. sativus | ||||
| A | B | A | B | A | B | A | B | |
| Control | 10.0 ± 0.0 | 0 | 9.7 ± 0.6 | 0 | 10.0 ± 2.0 | 0 | 8.7 ± 0.6 | 0 |
| 125 | 7.0 ± 1.0 * | 30.0 | 3.3 ± 0.6 **** | 65.9 | 3.0 ± 1.0 * | 70.0 | 9.7 ± 0.6 | −111.5 |
| 250 | 6.3 ± 1.5 ** | 37.0 | 2.0 ± 1.0 **** | 70.3 | 1.7 ± 1.2 ** | 83.0 | 9.3 ± 1.2 | −106.0 |
| 500 | 1.7 ± 1.5 **** | 83.0 | 10.0 ± 0.0 | −103.1 | 0.0 ± 0.0 *** | 100.0 | 7.0 ± 2.0 | 19.5 |
| 1000 | 0.0 ± 0.0 **** | 100.0 | 10.0 ± 0.0 | −103.1 | 0.0 ± 0.0 *** | 100.0 | 4.0 ± 1.0 ** | 54.0 |
| Radical Length (cm—A) and Percent Inhibition (B) | ||||||||
| µg/mL | L. multiflorum | S. arvensis | R. sativus | C. sativus | ||||
| A | B | A | B | A | B | A | B | |
| Control | 0.6 ± 0.4 | 0 | 0.8 ± 0.6 | 0 | 0.9 ± 0.3 | 0 | 1.0 ± 0.4 | 0 |
| 125 | 0.4 ± 0.2 | 33.3 | 0.2 ± 0.1 | 25.0 | 0.7 ± 0.0 | 22.2 | 2.3 ± 0.5 * | −230.0 |
| 250 | 0.3 ± 0.1 | 50.0 | 0.2 ± 0.0 | 25.0 | 0.0 ± 0.0 **** | 100.0 | 1.5 ± 0.4 | −150.0 |
| 500 | 0.0 ± 0.0 * | 100.0 | 0.0 ± 0.0 * | 100.0 | 0.0 ± 0.0 **** | 100.0 | 1.6 ± 0.5 | −160.0 |
| 1000 | 0.0 ± 0.0 * | 100.0 | 0.0 ± 0.0 * | 100.0 | 0.0 ± 0.0 **** | 100.0 | 0.2 ± 0.1 | 80.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polito, F.; Papaianni, M.; Woo, S.L.; Malaspina, P.; Cornara, L.; De Feo, V. Artemisia arborescens (Vaill.) L.: Micromorphology, Essential Oil Composition, and Its Potential as an Alternative Biocontrol Product. Plants 2024, 13, 825. https://doi.org/10.3390/plants13060825
Polito F, Papaianni M, Woo SL, Malaspina P, Cornara L, De Feo V. Artemisia arborescens (Vaill.) L.: Micromorphology, Essential Oil Composition, and Its Potential as an Alternative Biocontrol Product. Plants. 2024; 13(6):825. https://doi.org/10.3390/plants13060825
Chicago/Turabian StylePolito, Flavio, Marina Papaianni, Sheridan Lois Woo, Paola Malaspina, Laura Cornara, and Vincenzo De Feo. 2024. "Artemisia arborescens (Vaill.) L.: Micromorphology, Essential Oil Composition, and Its Potential as an Alternative Biocontrol Product" Plants 13, no. 6: 825. https://doi.org/10.3390/plants13060825