

Reproductive Capacity and Scanning Electron Microscopy (SEM) Analyses of the Micromorphological Surfaces of Three Endemic Satureja Species from Bulgaria
 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
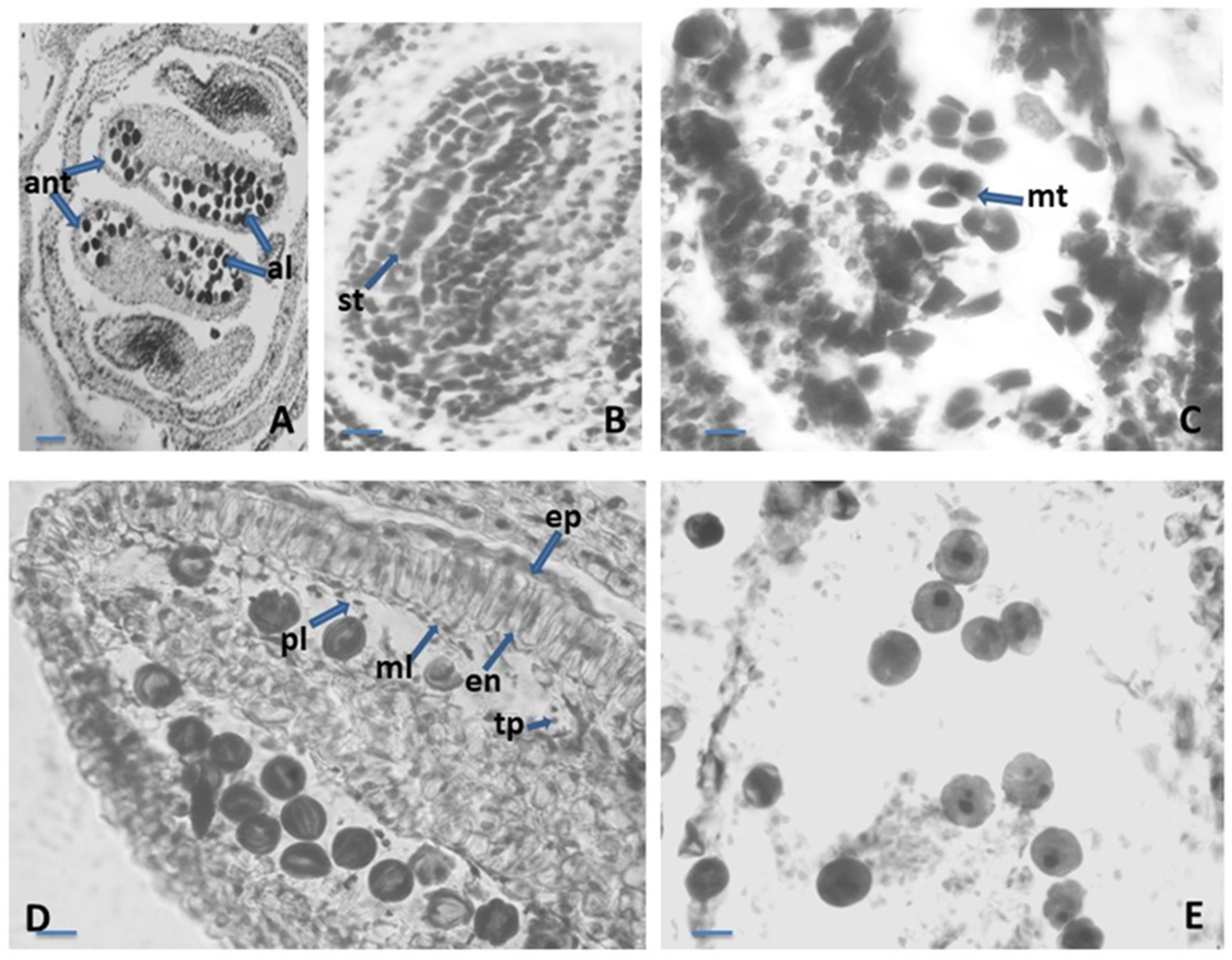
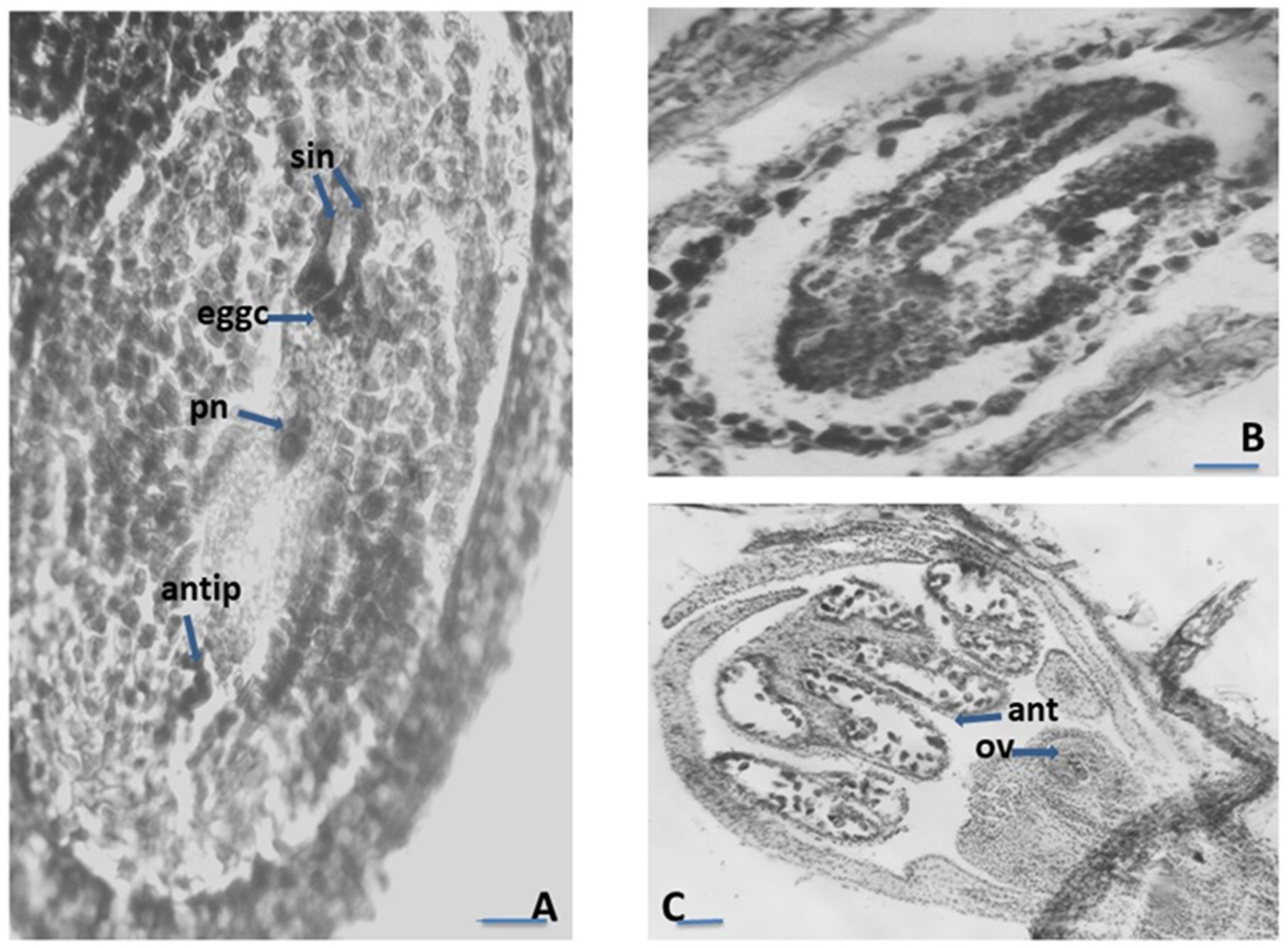
2.1. Embryological Analyses
2.1.1. Anther and Development of the Male Gametophyte
2.1.2. Ovule and Development of the Female Gametophyte
2.1.3. Nutlets (Seed) and Pollen Viability Testing
2.1.4. Nutlet (Seed) Germination
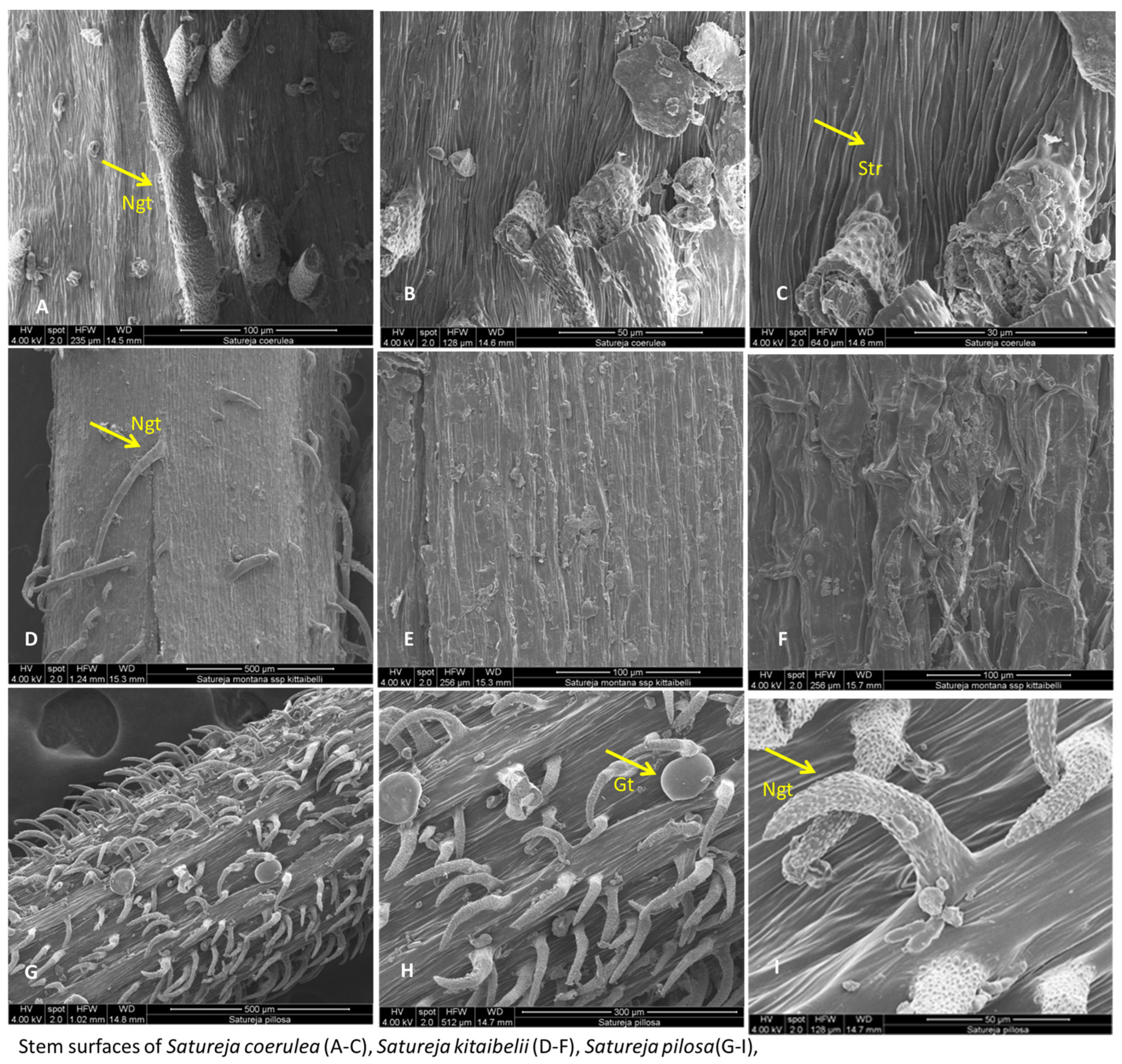
2.2. Scanning Electron Microscopy (SEM) Analyses
2.2.1. Leaves, Stem, and Calyx Surfaces
Satureja coerulea
Satureja kitaibelii
Satureja pilosa
2.2.2. Nutlet (Seed) Surfaces
Satureja coerulea
Satureja kitaibelii
Satureja pilosa
3. Discussion
3.1. Embryological Analyses
3.2. Scanning Electron Microscopy (SEM) Analysis
3.2.1. Leaves, Stem, and Calyx Surfaces
3.2.2. Nutlet (Seed) Surfaces
4. Materials and Methods
4.1. Collection of the Plant Materials
4.2. Embryological Analyses
4.2.1. Pollen Viability
4.2.2. Nutlet (Seed) Viability Testing
4.2.3. Testing of Nutlet (Seed) Germination under Different Light Regimes
4.3. Scanning Electron Microscopy (SEM) Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doroszenko, A. Taxonomic Studies of Satureja Complex (Labiate). Ph.D. Thesis, Edinburgh University and Royal Botanic Garden, Edinburgh, UK, 1986. Available online: https://www.researchgate.net/publication/319540144_Taxonomic_studies_on_the_satureja_complex_Labiatae (accessed on 14 April 2023).
- Başer, K.H.C.; Özek, T.; Kirimer, N.; Tümen, G. A Comparative study of the essential oils of wild and cultivated Satureja hortensis L. J. Essent. Oil Res. 2004, 16, 422–424. [Google Scholar] [CrossRef]
- Nooshkam, H.M.; Hadian, J.; Alemardan, A.; Mohammad, R.M. Drug yield and essential oil and carvacrol contents of two species of Satureja (S. khuzistanica Jamzad and S. rechingeri Jamzad) cultivated in two different locations. J. Appl. Res. Med. Arom. Plants 2017, 6, 126–130. [Google Scholar] [CrossRef]
- Ćavar, S.; Maksimović, M.; Šolić, M.E.; Jerković-Mujkić, A.; Bešta, R. Chemical composition and antioxidant and antimicrobial activity of two Satureja essential oils. Food Chem. 2008, 111, 648–653. [Google Scholar] [CrossRef]
- Skočibušić, M.; Bezić, N.; Dunkić, V. Phytochemical composition and antimicrobial activities of the essential oils from Satureja subspicata Vis. growing in Croatia. Food Chem. 2006, 96, 20–28. [Google Scholar] [CrossRef]
- Semerdjieva, I.B.; Zheljazkov, V.; Cantrell, C.L.; Astatkie, T.; Ali, A. Essential oil yield and composition of the Balkan endemic Satureja pilosa Velen. (Lamiaceae). Molecules 2020, 25, 827. [Google Scholar] [CrossRef] [Green Version]
- Zheljazkov, V.D.; Semerdjieva, I.B.; Cantrell, C.L.; Astatkie, T.; Aćimović, M. Phytochemical variability of essential oils of two Balkan endemic species: Satureja pilosa Velen. and S. kitaibelii Wierzb. ex Heuff. (Lamiaceae). Molecules 2022, 27, 3153. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Mooze, D.M.; Valeutine, D.H.; Walters, S.M.; Webb, D.A. Genus Satureja. In Flora Europaea; Tutin, T.G., Ed.; Cambridge University Press: Cambridge, MA, USA, 1972; Volume 3, pp. 108–124. [Google Scholar]
- Anchev, M. Genus Satureja. In Flora of Bulgaria; Yordanov, D., Ed.; BAS: Sofia, Bulgaria, 1989; Volume 9, pp. 335–342. (In Bulgarian) [Google Scholar]
- Šućur, J.; Popović, A.; Petrović, M.; Anačkov, G.T.; Bursić, V.; Kiprovski, B.; Prvulović, D. Allelopathic effects and insecticidal activity of aqueous extracts of Satureja montana L. J. Serbian Chem. Soc. 2015, 80, 475–484. [Google Scholar] [CrossRef]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic plant species conservation: Biotechnological approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef] [Green Version]
- Yıldız, F.; Aslay, M.; Kandemir, A.; Kaya, O. Reproductive biology of Fritillaria aurea Schott (Liliaceae), a rare species endemic to Turkey. Diversity 2022, 14, 1052. [Google Scholar] [CrossRef]
- Gopalakrishnan, K.K.; Thomas, T.D. Reproductive biology of Pittosporum dasycaulon Miq., (Family Pittosporaceae) a rare medicinal tree endemic to Western Ghats. Bot. Stud. 2014, 55, 15. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Lehman, C. Human-caused environmental change: Impacts on plant diversity and evolution. Proc. Natl. Acad. Sci. USA. 2001, 98, 5433–5440. [Google Scholar] [CrossRef] [Green Version]
- Murray, B.R.; Thrall, P.H.; Gill, A.M.; Nicotra, A.B. How plant life-history and ecological traits relate to species rarity and commonness at varying spatial scales. Austral Ecol. 2002, 27, 291–310. [Google Scholar] [CrossRef]
- Castro, S.; Silveira, P.; Navarro, L. How flower biology and breeding system affect the reproductive success of the narrow endemic Polygala vayredae Costa (Polygalaceae). Bot. J. Linnean Soc. 2008, 157, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.J. Systematics and reproductive biology. In Experimental and Molecular Approaches to Plant Systematics; Hoch, P.C., Stephenson, A.G., Eds.; Monogr Syst. Bot 53; Missouri Botanical Garden: St. Louis, MI, USA, 1995; pp. 263–272. [Google Scholar]
- Bernardello, G.; Anderson, G.J.; Lopez, S.P.; Cleland, M.A.; Stuessy, T.F.; Crawford, D.J. Reproductive biology of Lactoris fernandeziana (Lactoridaceae). Am. J. Bot. 1999, 86, 829–840. [Google Scholar] [CrossRef]
- Sharia, A.; Sefidkon, F. Investigation of pollination system and self-incompatibility intwo species of Satureja sahendica and S. bachtiarica. Iran. J. Rangelands Forest Plants 2020, 27, 204–215. [Google Scholar] [CrossRef]
- Daskalova, T.; Genova, E. Histological structure of the anthers and microsporogenesis in Hyssopus officinalis L. ssp. aristatus (Godr.) Briq. (Lamiaceae). Bot. Jahrb. Syst. 1996, 118, 297–302. [Google Scholar]
- Daskalova, T. Histological structure of the microsporangia, microsporogenesis and development of the male gametophyte in Nepeta cataria (Lamiaceae). Phytol. Balcan. 2004, 10, 241–246. Available online: http://www.bio.bas.bg/~phytolbalcan/PDF/10_2-3/10_2-3_18_Daskalova.pdf (accessed on 17 January 2023).
- Daskalova, T. On some specificities of seed formation in Salvia nemorosa (Lamiaceae). Phytol. Balcan 2004, 10, 7984. Available online: https://www.researchgate.net/publication/237790347 (accessed on 17 January 2023).
- Daskalova, T. Polygamy in some species of Nepeta (Lamiaceae). Phytol. Balcan. 2005, 11, 79–98. Available online: http://www.bio.bas.bg/~phytolbalcan/PDF/11_1/11_1_10_Daskalova.pdf (accessed on 17 January 2023).
- Daskalova, T. Polygamy in Lamium maculatum L. C. R. Acad. Bulg. Sci. 2008, 61, 327–334. Available online: https://www.researchgate.net/scientific-contributions/Tzvetana-Daskalova-2015103073 (accessed on 17 January 2023).
- Wightman, R. An overview of cryo-scanning electron microscopy techniques for plant imaging. Plants 2022, 11, 1113. [Google Scholar] [CrossRef] [PubMed]
- Husain, S.; Marin, P.D.; Šilić, Č.; Qaiser, M.; Petrović, B. A micromorphological study of some representative genera in the tribe Saturejeae (Lamiaceae). Bot. J. Lin. Soc. 1990, 103, 59–80. [Google Scholar] [CrossRef]
- Satil, F.; Tümen, G.; Akçelik, A.; Baser, K.H.C. Comparative morphological, anatomical, ecological and chemical studies on endemic Satureja parnassica subsp. sipylea from Turkey. Acta Bot. Croatica 2002, 61, 207–220. Available online: https://hrcak.srce.hr/3459 (accessed on 18 January 2023).
- Marin, M.; Ascensao, L.; Lakušić, B. Trichomes of Satureja horvatii Šilić (Lamiaceae): Micromorphology and histochemistry. Arch. Biol. Sci. 2012, 64, 995–1000. [Google Scholar] [CrossRef]
- Malmir, M.; Serrano, R.; Reza Gohari, A.; Silva, O. Characterization of Satureja khuzestanica Leaf as a herbal medicine. Microsc. Microanal. 2014, 20, 1425–1435. [Google Scholar] [CrossRef]
- Dodoš, T.; Janković, S.; Marin, P.D.; Rajčević, N. Essential oil composition and micromorphological traits of Satureja montana L., S. subspicata Bartel Ex Vis., and S. kitaibelii Wierzb. Ex Heuff. Plant Organs. Plants 2021, 10, 511. [Google Scholar] [CrossRef]
- Bräuchler, C.; Meimberg, H.; Heubl, G. Molecular phylogeny of Menthinae (Lamiaceae, Nepetoideae, Mentheae)—Taxonomy, biogeography and conflicts. Mol. Phylogen. Evol. 2010, 55, 501–523. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Salehi-Arjmand, H.; Movahedi, K.; Hadian, J. Molecular and morphological variability of Satureja bachtiarica in Iran. Plant Syst. Evol. 2015, 301, 77–93. [Google Scholar] [CrossRef]
- Hedge, I.C.; Feinbrun, N. Satureia thymbrifolia Hedge & Feinbrun sp. nov. Israel. J. Bot. 1969, 17, 213. Available online: http://www.worldfloraonline.org/taxon/wfo-0000304558 (accessed on 14 May 2023).
- Davis, G.L. Systematic Embryology of the Angiosperms; John and Wiley and Sons: Hoboken, NJ, USA, 1966. [Google Scholar]
- Johansen, D.A. A critical survey of the present status of plant embryology. Bot. Rev. 1945, 11, 87–107. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Embryology; Chronica Botanica: Waltham, MA, USA, 1950. [Google Scholar]
- Copeland, L.O.; Mc Donald, M.B. Principles of Seed Science and Thechnology, 4th ed.; Kluwer Academic Publishers: Hingham, MA, USA, 2001. [Google Scholar] [CrossRef]
- Peters, J. Tetrazolium testing handbook. In Contribution No 29 to the Handbook on Seed Testing Revised; The Association of Official Seed Analysts (AOSA); PMB: Las Cruces, NM, USA, 2000. [Google Scholar]
- Moore, R.P. Handbook on Tetrazolium Testing; International Seed Testing Association: Zürich, Switzerland, 1985; p. 99. [Google Scholar]
- Poddubnaya-Arnoldi, V.A. Characteristics of Flowering Plants Families According to the Cytoembryological Features; Nauka: Moscow, Russia, 1982. (In Russian) [Google Scholar]
- Kamellina, O.; Dzevaltovsky, A. Lamiaceae. In Comparative Embryology of Flowering Plants; Batygina, M., Yakovlev, T.B., Eds.; Nauka: Leningrad, Russia, 1987; pp. 225–236. (In Russian) [Google Scholar]
- Kai-yu, P.; Wen Jie, W.; Shi-liang, Z. Embryological study on Mosla chinensis (Lamiaceae). Acta Bot. Sinica 1997, 39, 111–116. Available online: https://www.jipb.net/EN/abstract/abstract26600.shtml (accessed on 18 January 2023).
- Jafari Marandi, S.; Arbabian, S.; Musavi, M. Structure and development of male and female gametophyte in Ocimum basilicum L. Caryologia 2017, 70, 102–111. [Google Scholar] [CrossRef]
- Yankova-Tsvetkova, E.; Yurukova-Grancharova, P.; Vitkova, A. Reproductive biology of the Balkan endemic Sideritis scardica (Lamiaceae). Bot. Serbica 2013, 37, 83–87. Available online: https://botanicaserbica.bio.bg.ac.rs/arhiva/pdf/2013_37_1_580_full.pdf (accessed on 17 January 2023).
- Zheljazkov, V.D.; Semerdjieva, I.B.; Stevens, J.F.; Wu, W.; Cantrell, C.L.; Yankova-Tsvetkova, E.; Koleva-Valkova, L.H.; Stoyanova, A.; Astatkie, T. Phytochemical investigation and reproductive capacity of the Bulgarian endemic plant species. Marrubium friwaldskyanum Boiss. (Lamiaceae). Plants 2022, 11, 114. [Google Scholar] [CrossRef]
- Aizen, M.A.; Ashworth, L.; Galetto, L. Reproductive success in fragmented habitats: Do compatibility systems and pollination specialization matter? J. Veg. Sci. 2002, 13, 885–892. Available online: https://www.jstor.org/stable/3236934 (accessed on 18 January 2023). [CrossRef]
- Aizen, M.A.; Feinsinger, P. Habitat fragmentation, pollination, and plant reproduction in a Chaco dry forest. Ecology 1994, 75, 330–351. Available online: https://www.jstor.org/stable/pdf/1939538.pdf (accessed on 25 May 2023). [CrossRef]
- Buchmann, S.L.; Nabhan, G.P. The Forgotten Pollinators; Island Press: Washington, DC, USA, 1996. [Google Scholar]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Ann. Rev. Ecol. Syst. 1998, 29, 83. [Google Scholar] [CrossRef]
- Gul, S.; Ahmad, M.; Zafar, M.; Bahadur, S.; Celep, F.; Sultana, S.; Begum, N.; Hanif, U.; Zaman, W.; Shuaib, M.; et al. Taxonomic significance of foliar epidermal morphology in Lamiaceae from Pakistan. Mycrosc. Res. Techniq. 2019, 82, 1507–1528. [Google Scholar] [CrossRef]
- Zaman, W.; Fazal Ullah, F.; Parmar, G.; Saqib, S.; Ayaz, A.; Park, S.J. Foliar micromorphology of selected medicinal Lamiaceae taxa and their taxonomic implication using scanning electron microscopy. Mycrosc. Res. Techniq. 2022, 85, 3217–3236. [Google Scholar] [CrossRef]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons, Systematic Anatomy of the Leaf and Stem, 2nd ed.; Claredon Press: Oxford, UK, 1979; Volume 1. [Google Scholar]
- Barthlott, W. Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nord. J. Bot. 1981, 1, 345–355. [Google Scholar] [CrossRef]
- Satil, F.; Kaya, A. Leaf anatomy and hairs of Turkish Satureja L. (Lamiaceae). Acta Biol. Cracov. Ser. Bot. 2007, 49, 67–76. Available online: https://abcbot.pl/pdf/49_1/09satil.pdf (accessed on 20 April 2023).
- Redžić, S.; Tuka, M.; Pajević, A. Research into microscopic structure and essential oils of endemic medicinal plant species Satureja subspicata Bartl. Ex Vis. (Lamiaceae). Bosn. J. Basic Med. Sci. 2006, 6, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunkić, V.; Bezić, N.; Ljubešić, N.; Bočina, I. Glandular hair ultrastructure and essential oils in Satureja subspicata Vis. ssp. subspicata and ssp. liburnica Šilić. Acta Biol. Cracov. 2007, 49, 45–51. Available online: https://abcbot.pl/pdf/49_2/06_dunkic.pdf (accessed on 20 April 2023).
- McDonald, I.; Kwong, M.B.I.; Francis, Y. Flowers-Seeds. In Flower Seeds: Biology and Technology; McDonald, I., Kwong, F., Eds.; CAB International: Wallingford, UK, 2005; p. 372. Available online: https://www.scribd.com/document/239950465/Flower-Seeds-Biology-and-Technology (accessed on 18 January 2023).
- Duletiæ-Lauševiæ, S.; Marin, P.D. Pericarp structure and myxocarpy in selected genera of Nepetoideae (Lamiaceae). Nord. J. Bot. 1999, 19, 435–446. [Google Scholar] [CrossRef]
- Kaya, A.; Satil, F.; Gogel, F. Nutlet surface micromorphology of Turkish Satureja (Lamiaceae). Biologia 2009, 64, 902–907. [Google Scholar] [CrossRef] [Green Version]
- Sundara, R.S. Practical Manual of Plant Anatomy and Embryology; Anmol Publications Pvt Ltd.: New Delhi, India, 2000; Available online: https://www.bagchee.com/books/BB28913/practical-manual-of-plant-anatomy-and-embryology (accessed on 18 January 2023).
- Heslop-Harrison, J.S. Pollen Capture Adhesion and Hydration. In Sexual Plant Reproduction; Cresti, M., Tizzi, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 81–88. Available online: https://link.springer.com/chapter/10.1007/978-3-642-77677-9_8 (accessed on 18 January 2023).
- Barthlott, W.; Ehler, N. Raster-Elektronenmikroskopie der Epidermis-Oberfl€achen von Spermatophyten. Trop. Subtrop. Pflanzenwelt 19; Akad. Wiss. Lit. Mainz; Franz Steiner Verlag: Stuttgart, Germany, 1977; p. 105. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variants | Satureja coerulea | Satureja pilosa | ||
|---|---|---|---|---|
| GE % ± SD | G % ± SD | GE % ± SD | G % ± SD | |
| V1 (N) | 24 ± 1.7 | 40 ± 4.0 | 21 ± 1.5 | 39 ± 2.5 |
| V2 (WR7:B1) | 20 ± 2.0 | 30 ± 2.0 | 9 ± 0.6 | 18 ± 1.0 |
| V3 (WR4:B1) | 31 ± 0.6 | 33 ± 1.5 | 13 ± 0.6 | 26 ± 1.7 |
| V4 (W) | 21 ± 1.5 | 30 ± 2.6 | 11 ± 1.5 | 23 ± 3.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semerdjieva, I.; Yankova-Tsvetkova, E.; Zheljazkov, V.D.; Koleva-Valkova, L.H.; Nikolova, R. Reproductive Capacity and Scanning Electron Microscopy (SEM) Analyses of the Micromorphological Surfaces of Three Endemic Satureja Species from Bulgaria. Plants 2023, 12, 2436. https://doi.org/10.3390/plants12132436
Semerdjieva I, Yankova-Tsvetkova E, Zheljazkov VD, Koleva-Valkova LH, Nikolova R. Reproductive Capacity and Scanning Electron Microscopy (SEM) Analyses of the Micromorphological Surfaces of Three Endemic Satureja Species from Bulgaria. Plants. 2023; 12(13):2436. https://doi.org/10.3390/plants12132436
Chicago/Turabian StyleSemerdjieva, Ivanka, Elina Yankova-Tsvetkova, Valtcho D. Zheljazkov, Lyubka H. Koleva-Valkova, and Rozalia Nikolova. 2023. "Reproductive Capacity and Scanning Electron Microscopy (SEM) Analyses of the Micromorphological Surfaces of Three Endemic Satureja Species from Bulgaria" Plants 12, no. 13: 2436. https://doi.org/10.3390/plants12132436