Figure 1. The Rise of Fishes 1995 book by fish expert John Long.
The majority of data sources
for fish here and in ReptileEvolution.com come from the excellent ink drawings of WK Gregory 1933 and the excellent photos of John Long 1995 in his book, ‘The Rise of Fishes’ (Fig. 1). Newer editions of the book are out there, but I don’t have them. Long’s own updates may erase some of the issues raised here.
Long 1995 writes:
“The first osteichthyans are very poorly known from fossils, represented by a few scales and a mere fragments of bone. The oldest articulated remains, showing what their bodies and heads were like, are about 400 million years old.”
The LRT demonstrates
restricting phylogenetic fish taxa to just Silurian and Devonian fossils is unnecessary and restrictive. There are plenty of extant, yet still primitive, taxa one can plug into any fish phylogenetic analysis, like the large reptile tree (LRT, 1656+ taxa), for one. Without fossils the LRT can recreate the fish ‘tree of life’ starting with lancelets, then sturgeons, then sharks, and ending with sea horses, anglerfish and tetrapods.
Long includes sturgeons in the clade of bony fish (Osteichthyes)
despite his notes that sturgeon skeletons retain large amount of cartilage and much of the external armor is bone. In the LRT jawless armored Osteostraci are basal to sturgeons prior to the evolution of terminal jaws.
Long “pulls a Larry Martin”
when he states, “Osteichyan fishes are characterized by having a well-ossified internal bony skeleton” then backtracks by noting, “although the earliest fossil forms show the least degree of ossification of the vertebrae and internal bones.”
By contrast,
in the LRT only the last common ancestor of all extant bony fish (sans sturgeons and paddle bills) determines clade membership, despite the presence or absence to certain ‘key’ traits, like a bony internal skeleton.
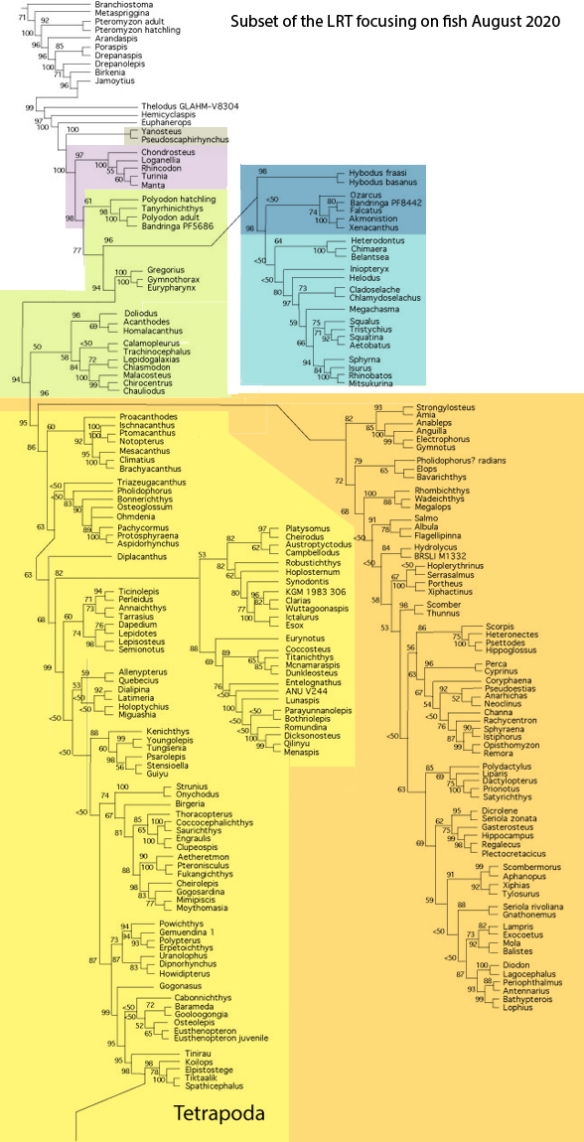
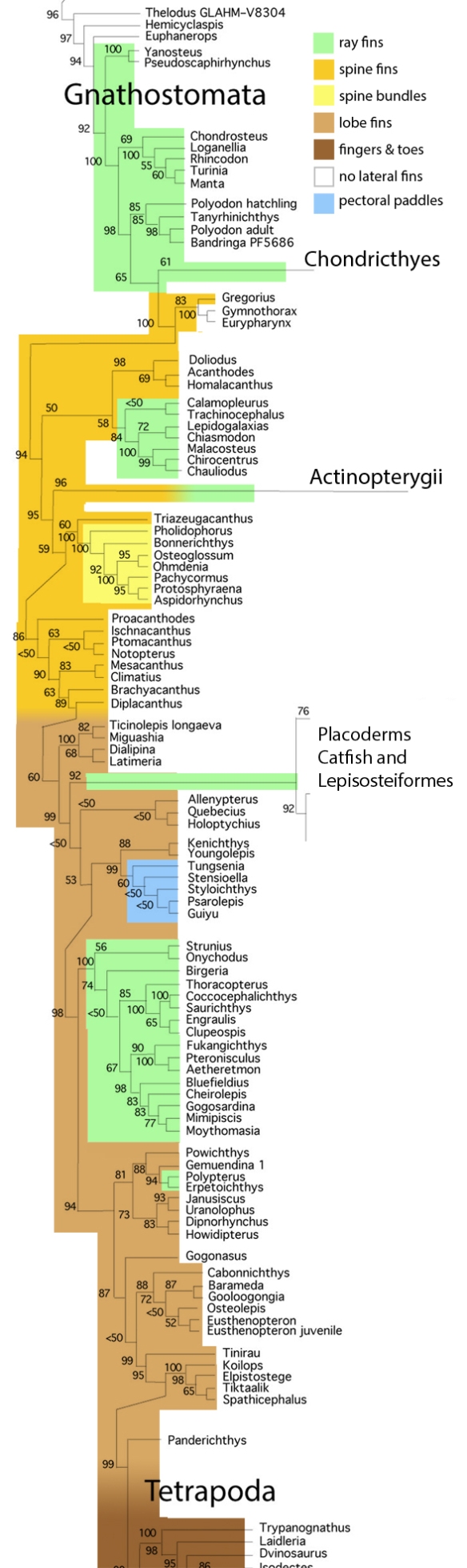
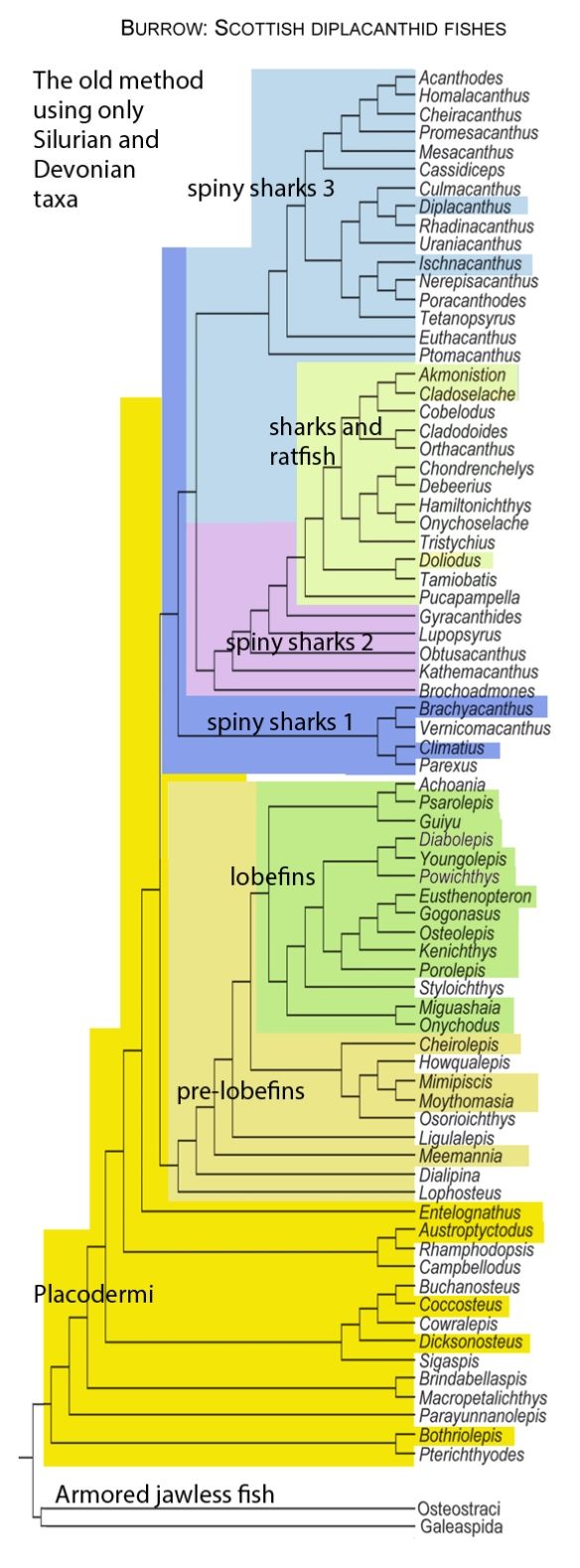
Figure x. Subset of the LRT focusing on fish.
The LRT confirms Long’s 1995 fish cladogram
with regard to nesting jawless fish (Agnatha) basal to jawed fish (Gnathostomata), naturally.
In Long 1995, Osteostraci
is the proximal outgroup to the Gnathostomata.
In the LRT, a member of the Osteostraci is basal to tube-mouth sturgeons in the LRT. Members of the Thelodonti are basal to Gnathostomata in the LRT.
In Long 1995, the basal dichotomy in Gnathostomata
splits Placodermi from Acanthodii. Those two are not related in the LRT.
In the LRT basal Gnathostomata splits taxa with transverse toothless jaws (Loganellia + Rhincodon + Manta) from taxa with U-shaped toothy jaws (Falcatus) in the LRT. No suprageneric taxa are employed (exception: Reptilia, Lepidosauromorpha, Archosauromorpha).
In Long 1995, Silurian Placodermi give rise to
“protosharks”, then Cladoselache, then Holcephalomorpha (ratfish) and Neoselachii (true sharks)
In the LRT, Placodermi include and give rise to catfish, all derived from Cheirodus/Amphicentrum. In the LRT, the second great dichotomy splits ratfish + sharks apart from xenacanthid ‘sharks‘, hybodontid ‘sharks‘ and Pachycormus.
In Long 1995, Silurian Acanthodii give rise to
Lophosteiformes (known only from ornate epidermal scales) and Actinopterygii (ray-finned fish)
In the LRT, Acanthodii are not primitive, but arise from ray-fin fish like the Cretaceous, bony-finned Bonnerichthys and the extant Osteoglossum.
Cause for concern
This major time gap between known fossils and phylogenic first appearance is cause for concern on the one hand, and a call to action on the other. At present it does not make sense that placoderms and acanthodians (spiny sharks) are present in the Silurian while intervening and more primitive taxa in the LRT are no yet known from the Silurian… and yet, we’ve seen this before with Jurassic multituberculates preceding more basal members of the placental clades, Glires, Carnivora and Primates.
Phenomic phylogenetic analyses, like the LRT, deliver
the gradual accumulation of derived traits that support evolutionary theory. Missing Silurian taxa will have to be discovered whenever someone discovers them.
On the other hand,
Long 1995 makes no attempt to provide any generic last common ancestors with jaws for Acanthodii + Placoderm, nor does he spell out any gradual accumulations of derived traits at transitional points, not does he employ more than a token number of generic or specific taxa in his cladogram. Suprageneric taxa are often a problem in taxonomy, as noted earlier. They can be too easily cherry-picked.

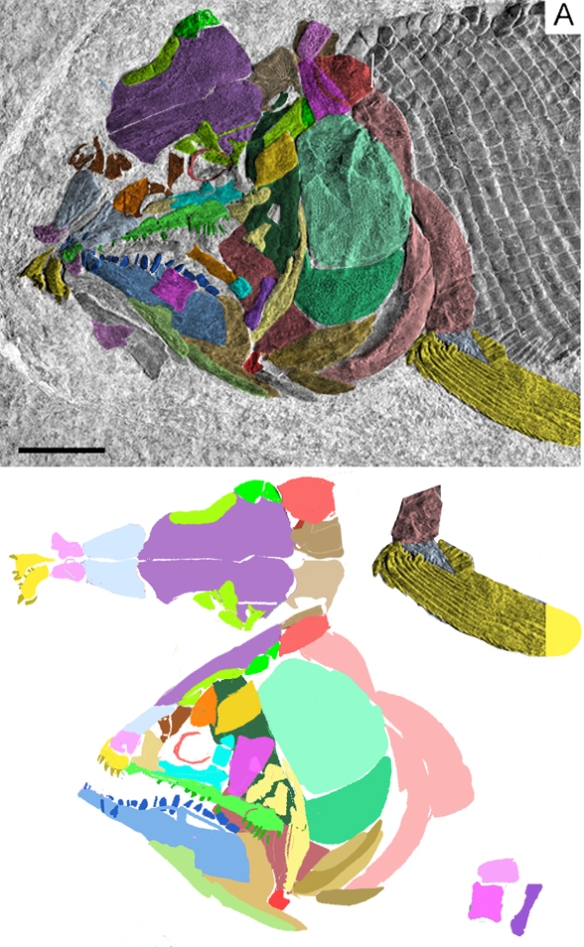
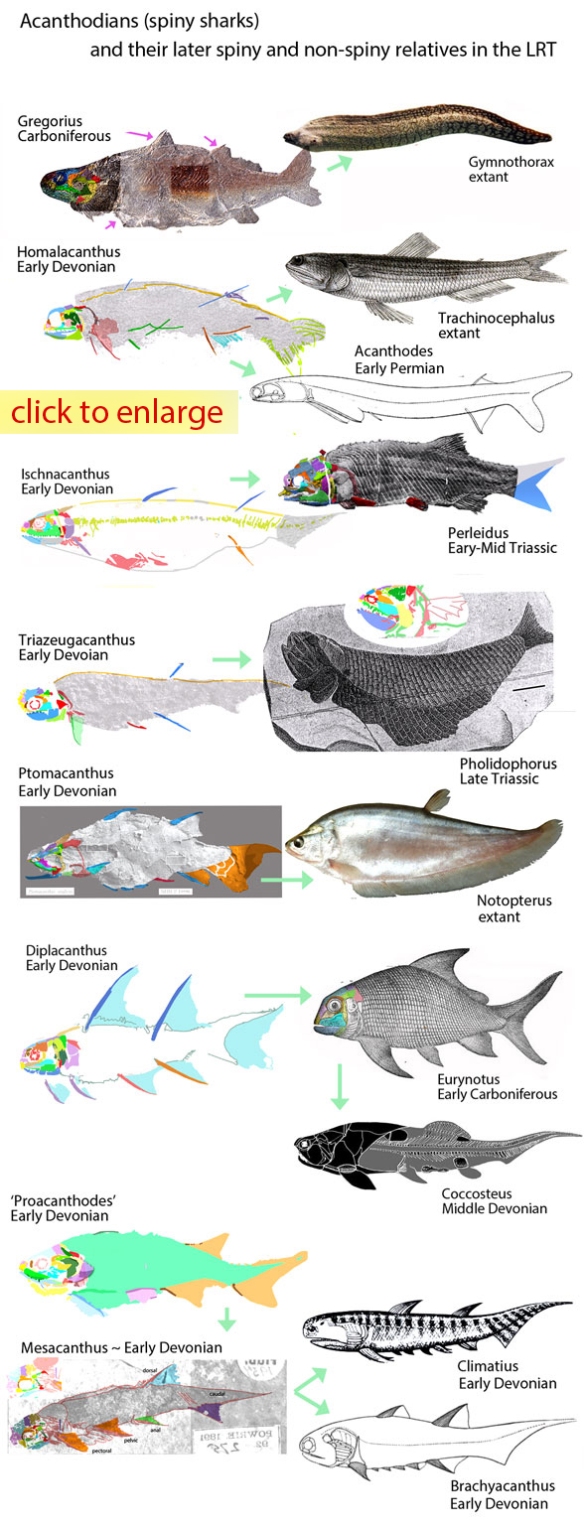
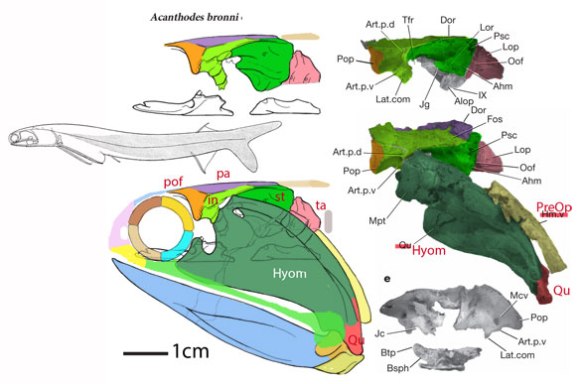
Figure 2. Early Devoniann Mesacanthus in situ. This 3 cm fish is a typical acanthodian here traced using DGS methods and reconstructed. Distinct from other spiny sharks, this one lacks large cheek plates, as in the extant Notopterus (Fig. 3).
So, are placoderms and acanthodians ‘bony fish’?
In the LRT: yes, despite having less than fully ossified internal skeletons. That could be a retained primitive trait or a reversal in both cases. Both are derived from taxa in which the external scales are robust, providing support to the body, allowing the skeleton to degenerate. As for that heterocercal tail in acanthodians (Fig. 2) and placoderms… based on the evidence, that is a primitive trait, not a reversal. Intervening taxa, like extant Trachinocephalus, appear to have independently evolved a diphycercal tail and bony skeleton, which their Silurian direct ancestors probably lacked.
Cherry picking traits,
even ‘important’ traits, is ‘Pulling a Larry Martin‘, something the wise professor taught us not to do. Find your clades using specific or generic taxa tested against 200+ traits to find that last common ancestor, no matter the ‘key’ traits that tickle your fancy.
References
Gregory WK 1933. Fish skulls. A study of the evolution of natural mechanisms. American Philosophical Society 23(2) 1–481.
Long JA 1995. The Rise of Fishes. Johns Hopkins University Press. Baltimore and London.